الإدراك في أصغرِ الخلايا | دانيال دينت ومايكل ليفين – ترجمة: حيَّان الغربي
Dec 5, 2023 تستغرق 10 دقائق للقراءة

تحرير ومراجعة: بلقيس الأنصاري
الأفقُ الكبير المُقبِل لعِلم الأحياء: فهم الخلايا والأنسجة والكائنات الحيَّة بوصفها وسائط لها جداول أعمالها الخاصة، (وإن لم تكن موجَّهةً استنادًا إلى الأفكار).
يطيب لعلماء الأحياء النظر إلى أنفسهم بوصفهم سلوكيين عِلميين يفسِّرون ويتكهّنون بالطرق التي تسلكها البروتينات والعضلات والخلايا والنباتات والحيوانات وسائر الأحياء ضمن ظروفٍ معينة، وذلك بفضل أجزائها الصغرى. وهُم يحددون آلياتٍ سببية تنفّذ وظائف متنوّعة تنفيذًا موثوقًا من قبيل نسخ الحمض النووي الريبوزي منقوص الأكسجين- دي.إن.إيه، ومهاجمة المستضدات والتركيب الضوئي، وتقدير درجات الحرارة، وإمساك الفريسة، وإيجاد طريق العودة إلى المأوى وهلمّ جرًّا، بيدَ أنهم لا يعتقدون أنَّ هذا الإقرار بالوظائف يورّطهم في غايةٍ مذمومة، أو تنسيب للأسباب والأغراض، أو فهم للخلايا والأجزاء الأخرى من الآليات التي يتحرّونها.
ولكن حين أدارت “العلوم الاستعرافية” -علوم الإدراك- ظهرها للسلوكية منذ ما يربو على 50 عامًا، وشرَعت في التعامل مع الإشارات والخراط الداخلية والأهداف والتوقعات والمعتقدات والرغبات، تعرَّضت صفوف علماء الأحياء للخلخلة والاهتزاز. فلا شكّ أنهم سلَّموا بأنَّ البشر وبعض الحيوانات يملكون عقولًا، وأنَّ أدمغتهم تجسِّد العقول المادية -بدلًا من كونها عقولًا ثانوية مبهمة، تعالج المعلومات، ويستهدي بها السلوك الموجّه. وأمَّا الحيوانات التي لا أدمغة لها كالكيسيات، فإنها لا تمتلك عقولًا، وهو ما ينطبق أيضًا على النباتات والفطريات والجراثيم. فهم قد قاوموا اعتماد مصطلحات مفتعلة ضمن عملهم التنظيري إلاّ بوصفها مجازًا مفيدًا عند التعليم أو شرح الأفكار لجمهورٍ من غير المختصين، ومن ثَمَّ لم تكن الجينات تتسم بالأنانية حقًّا، ولم تبذل الأجسام المضادة مساعيَ حقيقيةً، ولم تدرك الخلايا مكان وجودها حقًّا، ولم تكن تلك الآليات البيولوجية الصغيرة وسائط تحمل جداول أعمالها الخاصة، وإن كان التفكير بأنها حقًّا وسائط لديها جداول أعمالها الخاصة غالبًا ما أفضى إلى إكسابنا المزيد من المعارف.
ونعتقد أنَّ هذا الاحتراس العِلمي المحمود قد مضى بعيدًا حتى اللحظة، وهذا يكبّل أيدي علماء الأحياء، ويحول بينهم وبين استكشاف أبرز الفرضيات المبشِّرة، تمامًا كما منعت السلوكية علماء النفس من النظر في كيفية التمكّن من تفسير سلوكيات مرضاهم القابلة للقياس بصفتها مؤثّرات الآمال والمعتقدات والخطط والمخاوف والنيّات والاضطرابات العقلية، وهلمّ جرًّا. وذات مرّة، طرح سيدني مورجنبيسر، الفيلسوف الفطن، على بي.إف سكينر السؤال التالي: “أتعتقد أنَّنا ينبغي ألا نؤنسِن البشر؟”. فيما نقول نحن إنه ينبغي لعلماء الأحياء التحلّي بالهدوء والنظر في فضائل أنسنَة جميع أنواع الكائنات الحيَّة. ففي نهاية المطاف، أليس علم الأحياء نوعًا من الهندسة العكسية لجميع أعضاء الكائنات الحيَّة وعملياتها؟ منذ التقدّم السيبراني الذي شهدته أربعينيات القرن المنصرم وخمسينياته، ألفى المهندسون أنفسهم إزاء علمٍ عمليّ راسخ يرتكز على آلياتٍ هادفة وموجَّهة لم تترك شيئًا للروحانيات والماورائيات؛ لذا نقترح نحن أن يدرك علماء الأحياء مَن سبقهم.
لعلَّنا نوافق على أنَّ الإسراف في نسبِ الغايات للأشياء يعدّ خطأ، فإذا كانت قوانين نيوتن قد اهتمَّت بتوقّع الطريق الذي ستسلكه الكُرة من أعلى التلّ، ولكنَّ هذه القوانين عينها تعجز عن مساعدتنا على فهم ما قد يفعل فأر عند قمة التلّ. لذا تتمثّل الطريقة الأخرى لارتكاب الخطأ في الفشل في اعتبار نظامٍ ما موجّهًا نحو هدفٍ محدد إذا كان حقًّا أيضًا، فهذا النوع من رهاب الخطط الغائيّة يُعيق بصورةٍ جوهرية المقدرة على توقّع الأنظمة المعقدة والسيطرة عليها ما دام أنه يحول دون اكتشاف ضوابطها الأكثر فعالية أم نقاط ضغطها الداخلية. ولعلَّنا نرفض الماهويّة البسيطة التي تفيد أنَّ للإنسان غاياتٍ “واقعية”، أمَّا كل ما عدَاه فله “ما يشبه” الغايات من الناحية المجازية فقط. ويظهر لنا التقدّم الأخير في المعرفة القاعدية والعلوم ذات الصِلة كيفية تجاوز هذا النوع من التفكير العدمي بشأن “الحيوان الإنسان”، من خلال تطبيع القدرات الإنسانية، ومقايضة تميّز مزدوج ساذج بسلسلةٍ متصلة لمقدار الوساطة التي تمارسها أيّ منظومة.
ويعود الفضل لتشارلز داروين، في إعفاء علم الأحياء تمامًا من الاضطرار إلى استحضار “مصممٍ ذكيّ”، ينسب إليه خلق جميع تلك الآليات؛ فما زال التطوّر بالاصطفاء الطبيعي يتولّى مسؤولية النهوض بكل ذلك العمل القائم على التنقية والتركيز والتمايز. إذ إننا جميعًا آليات جسدية ذات آلياتٍ فيزيائية تخضع لقوانين الفيزياء والكيمياء. بيدَ أنَّ ثمَّة فارقًا عميقًا بين الآليات الإبداعية الذكية التي يعدّها المصمم الإنسان الذكيّ، من قبيل ساعات الحائط والمحركات وأجهزة الكمبيوتر على سبيل المثال، والآليات المصممة والمركبة بواسطة الاصطفاء الطبيعي. ولعلَّنا قادرون على تحديد هذا الفارق بالاعتماد على شيءٍ يسير من المخيّلة.
تصوّر أنك قد اشتريت سيارةً يتم التحكّم بها عن بُعد، ثمّ وصلتك في صندوقٍ كبيرٍ كُتِبَ عليه: “هذا المنتج بحاجةٍ لبعض أعمال التجميع”. وبمجرّد أن تفتح الصندوق ستجد مئات القطع المختلفة التي تفتقر إلى بطاقات التعريف، وأنك لن تجد كتيّبًا للتعليمات لمساعدتك على تجميع تلك القطع أيضًا. ستجد نفسك في مواجهة مهمةٍ مهولةٍ إذن، ولا تتعلّق المشكلة بمدى خفّة بنانك وقوتها، إنما بعدم درايتك بكيفية التركيب أصلًا. وفي هذه الحالة لا شك أنك كنت تثمّن عاليًا صنيع من يزودك بدليل تركيب حسن الإعداد مع مخططات بيانية وبطاقات تعريف لسائر القطع، ولكنَّ هذا ما كان ليتمّ لولا قدرتك على رؤية المخططات وقراءة التعليمات والبطاقات. فلو زُوِّدت بدليلٍ باللغة الروسية، لكان الأمر بلا طائل إن كنت لا تجيد قراءة هذه اللغة. علاوةً على ذلك، يجب عليك أن تعرف كيف تركّب الظفر “أ” ضمن الفتحة “ب”، والبرغي رقم “17” في الصمولة رقم “95” مثلًا.
ولكن ها أنت تلمح قصاصةً ورق تفيدك أن تضع جميع القطع في إناءٍ كبيرٍ مملوءٍ بالماء، ثمّ تضعها على الموقد وتحرّكها حتى الغليان. وهكذا، تفعل أنت ما تطلبه منك الورقة لتذهلك القطع، وهي تلتحم ببضعها في كتلٍ صغيرةٍ لا تلبث أن تكبر شيئًا فشيئًا، وكل ظفرٍ يعثر على فتحته كما يدور كل برغيٍّ ضمن صمولته، ولا شيء يوجّه هذه القطع سوى الماء المغلي باهتياجه العشوائي. وفي بضع ساعات، ستكون مهمة تركيب سيارتك الجديدة قد تمَّت بنجاح، وليس عليك سوى أن تجفّفها من الماء قبل أن تشرع في قيادتها. يبدو أنَّ المخيّلة ذهبت بنا إلى المستحيل بالطبع، بيدَ أنها تردد صدى “معجزة” الحياة التي تأخذ قائمة القطع وكتيّب التعليمات الخاصَّيْنِ بالدي.إن.إيه، دون مساعدةٍ من “مجمِّعٍ ذكي”، وتتولّى تركيب كائنٍ جديدٍ من ملايين القطع المتحرّكة الموصولة ببعضها بصورةٍ صحيحة.
لدينا بالفعل سرديةً مفصّلةً برّاقةً حول كيفية قراءة قائمة المكوّنات واستيفائها، من الجينات إلى البروتينات، بفضل تلك الأجهزة الرائعة والجسيمات الريبية والشابرونات وغيرها، ثمّ إننا نحرز تقدّمًا بارزًا في فهمنا لكيفية إصدار الدي.إن.إيه لتعليمات التجميع. وما دام هذا المنحى البحثي التصاعدي قد حقَّق نجاحًا كبيرًا حتى اللحظة، أفلن يميط اللثام عن تفاصيل جميع العمليات التي تؤدي وظيفة إناء الماء الساخن الذي جادت به مخيّلتنا في نهاية المطاف؟ وهل هذه مشكلة حقًّا؟
نحن نعتقد أنها كذلك، فقد أُحرز التقدّم الكبير على صعيد التنقيب وصولًا إلى مستوى الجزيء بصورةٍ رئيسة، أمَّا على المستويات الأعلى، فليس التقدّم كبيرًا إلى هذه الدرجة بطبيعة الحال. فما زلنا ضعفاء نوعًا ما على صعيد التحكّم بالبنية التشريحية أو معرفة طريقة إعادتها إلى مسارها الصحيح في حالة السرطان، ولهذا ليس لدينا طب تجديدي حقيقي حتى الساعة. وإذا كنّا نعرف كيف نحدِّد مصائر الخلايا الفردية من خلال الخلايا الجذعية، فما زال علينا أن نقطع شوطًا كبيرًا قبل أن نتمكّن من صناعة أعضاءٍ معقدة غبّ الطلب. ولعلَّنا غير قادرين على صناعة تلك الأعضاء إلاّ في بضع حالاتٍ، وهي الحالات التي تعلَّمنا فيها التواصل مع مجموعة الخلايا التي توفّر منشطًا يسيرًا، من قبيل النمط الكهروحيويّ، الذي يصدر الأمر التالي، مثلًا: “ابنِ عينًا هنا”، ومن ثمّ نترك لذكاء مجموعة الخلايا تولّي المهمة الشاقة والتوقف لدى إنجاز العضوّ المطلوب.
كلَّما ازدادت مقدرة الوسيط على التأقلم مع تدخّلك؛ ارتفع مستوى الكفاءة التي يظهرها.
إذا ما التزمنا بالمقاربة التصاعدية، فإنَّ جميع الجهود التي نبذلها على صعيد الهندسة الحيويَّة ستتوقّف إثر قطاف الثمرة التي كانت في المتناول (المثانات المصنّعة بالطباعة ثلاثية الأبعاد وما شابهها). لعلَّ جيلنا لن يتمتع بالقدرة على الإدارة الدقيقة والمفصّلة لأنواع الخلايا وصولًا إلى تكوين يدٍ أو عينٍ بشريةٍ قادرةٍ على تأدية وظائفها، ولن تكون لدى مقاربات التحرير الجينومي أيَّة فكرة عمَّا ستحرّره الجينات لبناء الأعضاء المعقدة اللازمة للترميم أو الازدراع. التعامل مع الخلايا يبدو كأنه قطع من الطوب الأبكم التي يجب إدارتها بصورةٍ مفصّلة بمثابة ممارستنا للعبة ما بأيدٍ مكبّلة خلف ظهورنا بما يفضي إلى “شتاءٍ جينومي”، فيما لو توقّفنا عند هذا المستوى الجزيئي على وجه الحصر، فهذا ما يثبت لنا من خلال غياب التقدّم في التحكّم التخلّقي العقلاني. فمثلًا رغم سائر الأبحاث الجيدة في العلوم بشأن تنظيم الخلايا الجذعية المستندة إلى الدي.إن.إيه في المستورقات، إلّا أنَّ جميع النماذج البيولوجية الجزيئية المستخدمة في هذه الأبحاث لا يقدّم أيّ توقعٍ بشأن هذه التجربة الفكرية اليسيرة.
فلنأخذ دودة مسطحة الرأس ولنزرع فيها خلايا جذعية من ديدانٍ مثلثة الرأس بنسبة قدرها 50%، فإذا ما قمنا بقطع رأسها بعد اختلاط الخلايا الجذعية، ما الشكل الذي سنحصل عليه حين يتجدد الرأس؟ ما الذي سيحدث حين ترغب 50% من الخلايا الجذعية بتشكيل رأسٍ مستوٍ بينما ترغب النسبة المتبقية بتكوين رأسٍ مثلث؟ هل سيربح أحد الجانبين أم سيكون للرأس شكلٌ وسط ما بينهما؟ أو لعلَّ عملية التشكّل لن تنتهي ما دام بعض الخلايا لن يرضى بشكل الرأس الناجم على الإطلاق.
تعجز تفاصيل علم الأحياء الجزيئي عن دعم توقّع محدد؛ لأنها تتصدى للمسائل على المستوى الخلوي بينما لا تقارب الإشكالية المتعلّقة بكيفية تحديد المجموعة للشكل الكبير الذي ستبنيه وكيفية تحديدها لحظة التوقف حين يتم إنجاز هذا الشكل. وإنه لمن الجائز تجريبيًّا طرح السؤال بشأن تمثيل الأشكال التشريحية المستهدفة في النسج -وقد تم طرحه تجريبيًّا حقًّا- فقط عندما ينظر المرء نظرة جادَّة إلى أنَّ المجموعة لا تتمتع بمستوى تحليل آلي فحسب، بل بمستوى تحليل قائم على معالجة المعلومات أيضًا. وذلك أساس ما كانت عليه حال علوم الأعصاب حين اقترح ديفيد مار في كتابه البارز: “رؤية”، الصادر عام 1982، أنَّ التقدم يعتمد على تحرّي ما أسمَاه بـ “المستوى الحسابي”، وقد عنَى به المستوى الذي يحدد عنده المرء المهام الاستعرافية؛ كي يتمكّن من البحث عن السيرورات المعلوماتية المساعدة على تنفيذ تلك المهام.
ينطوي التفكير في مكوّنات الكائنات الحيَّة بوصفها وسائط تتحيّن الفرص ساعيةً إلى إنجاز مهام معينةٍ على المجازفة، بيد أنَّ المردود المعرفي الذي قد يعود به، قد يكون كبيرًا حقًّا. فلنفترض أنك تتدخل بالخليَّة أو التركيب الخلوي أثناء التطور، فتقوم بتحريكها أو انتزاعها من بين جيرانها المعتادين لتتبيّن هل هي قادرةً على التعافي والعودة إلى أداء دورها الطبيعي. هل تستطيع الخليَّة تحديد مكانها؟ هل ستحاول العثور على جيرانها؟ أو لعلَّها ستشرع في تنفيذ مهامها العادية حيثما هي الآن؟ أو لربما ستجد لنفسها عملًا آخر لتقوم به. كلّما ازدادت مقدرة الوسيط على التأقلم مع تدخّلك ارتفع مستوى الكفاءة التي يظهرها. وحين “ترتكب [الخليَّة] خطأً”، فما الذي يجعلها ترتكبه؟ أيمكنك أن “تخدع” تلك الخليَّة لتبكّر أو تتأخر في تصرفاتها؟ ولهذه التجارب على مستوى النسج والأعضاء آلاف التجارب التي تشبهها ضمن مجال العلوم الاستعرافية التي تفضي إلى أوهامٍ أو تشوّشاتٍ غريبةٍ أو عمىً محليّ بحثها علم الأمراض الذي يوفّر الأفكار حول كيفية إنجاز “السِحر”، ولكن فقط من خلال الحفاظ على مسارٍ يعرفه الوسيط ويريده.
وفيما يلي طريقة يسيرةٌ أخرى للتفكير في المشكلة، فبمجرد ولادة الخلايا المبكرة الفردية (الخلايا الجذعية على سبيل المثال)، من الواضح أنها تتولّى مسؤولية الاهتمام بتطوّرها الخاص، فتشرع بتشكيل نفسها والوسط المحيط بها في الآونة نفسها دون حاجةٍ لتعليماتٍ أخرى من والديها. وتصبح هذه الخلايا مستقلّة ذاتيًّا إلى حدٍّ ما بخلاف التروس والمكابس (المفتقرة للعقل) في محرّكٍ مصمّم بذكاء. تجد الخلايا طريقها، فما الذي يمكنه أن يفسِّر الأمر؟ أهو مسارٌ مماثل لمسارٍ محدد بفتات الخبز؟ نعم، في بعض الحالات، بيدَ أنه ينبغي أن تتحلَّى الخلايا بالذكاء الكافي لتحرّي المسار واقتفاء الفتات. لعلَّنا قد نأمل بتفسيرٍ فيزيائيّ يسير نسبيًّا.
فلندع المخيّلة تتفتّق عن صورةٍ أخرى. يستند الأمر برمته إلى قوة الجذب المغناطيسية، إذ تنتقل الأجزاء إلى مواطنها المناسبة من خلال آلاف المجموعات المتنوعة والمولّفة توليفًا مختلفة للجذب المغنطيسي. وقد أشار طاليس الفيلسوف الإغريقي إلى أنَّ حجر المغناطيس لديه “روح” ترغب ببرادة الحديد وتحتاج للاقتراب منها. لا شكّ أنه كان مخطئًا، ولكن هل لنا أن نفسِّر المساعي الحثيثة والرغبة القويّة البيّنة لدى الخلايا من خلال الاحتجاج بشيءٍ من هذه القوى الفيزيائية الانكماشية؟ لا يبدو أنَّ ثمَّة مجالًا لذلك؛ إذ لم يُكتشف أيّ من هذه القوى الفيزيائية الأساسية الموسومة بالتمييز المتعدد، ثمّ إنَّ ثمَّة أسبابًا نظريةً عميقةً تكمن وراء استنتاج أنه من المتعذر وجود أيّ قوى كهذه. ولكن ماذا عن المغناطيس الافتراضي؟
نستطيع الآن أن نصنع مغناطيسًا افتراضيًّا يجتذب أيّ شيئين مما قد يحلو لكم، فنستطيع صنعه من برمجيات الكمبيوتر. ولعلَّ طريقة التصاق مؤشّر الفأرة على الجزء المستهدف ضمن برنامج “وورد”، وطريقة التصاق لون الخلفية المستخدم للتشديد بعددٍ من الكلمات ضمن السطر؛ تجسّدان مثالين على مغناطيسين افتراضيين يَسِيرَيْن يعملان من خلال توزيع المعلومات.
وقد غدا وصف هذه الظاهرة بمساعدة الاصطلاحات المؤنسِنة على نحوٍ متعمّد ممارسةً معيارية، فحين ننقر بالفأرة، نأمر المؤشر بإمساك الشيء المستهدف على الشاشة، وحين نحرّك الفأرة ننقل الشيء المعني على الشاشة حتى نشير للمؤشر بإفلات ذلك الشيء من خلال النقر بالفأرة مجددًا. وقد خضع هذا الحديث حول إرسال الإشارات ومعالجة المعلومات للتبسيط بشكلٍ واضح بفضل أجهزة الكمبيوتر؛ حيث لا وجود لقوى الفيزياء المُبهمة. وقد قُوبل هذا الأمر مقابلة صحيحة بوصفه ترخيصًا لاعتماد هذا الحديث بشأن المعلومات في شتى أقسام علم الأحياء. وقد غدت الكشّافات والإشارات ودارات التغذية الراجعة وسيرورات صنع القرار لَبِنَات بناءٍ فيزيائي لا جدال عليها في علم الأحياء المعاصر، مثلما هو حالها في مجال الكمبيوتر. بيد أنَّ ثمَّة فارقًا ينبغي التطرّق إليه؛ إذ يفضي إخفاقنا في الانتباه له إلى تعويق مخيّلة المنظّرين. فبتعبير يستوجب منّا تحليله بعنايةٍ أقول: ليست الخلايا الفردية مجرّد لبنات بناءٍ فحسب -كحال الترس في الآلة أو المضخة- بل تتمتع الخلايا بكفاءاتٍ أخرى تجعل منها وسائط (غير موجهة) تعتمد على المعلومات التي تحملها في طياتها للمساعدة على تجميع نفسها ضمن بنًى أكبر، وفي مشاريع أخرى واسعة النطاق مما لا تحتاج إلى فهمه.
ويميل “الإنسان العاقل” Homo sapiens”، إلى التسليم جدلًا بمَلَكَات الهندسة، فعلى مرّ آلاف السنين تطلّع أسلافنا إلى الأنساق الفيزيائية التي ألفوا في أنفسهم القدرة على استغلالها من خلال تصميم البنى والهياكل القادرة على تنفيذ موثوق لوظائف معينة. فما الذي يصنع حبلًا جيدًا أو غراءً جيدًا أو مشعلًا جيدًا للنار؟ فالبرغي والصمولة كوسيلةٍ للتثبيت يجسّدان استغلالًا مصمَّمًا بأناقةٍ للعتل والمرونة وقوة الشد والاحتكاك، وقد تطوّرت هذه الأداة على مدى ألفي عام، وتعرّضت لتعديلاتٍ جوهريةٍ خلال المئتي سنة المنصرمة. ما زال التطوّر بـ “الاصطفاء الطبيعي” يضطلع بالتطلّع ذاته على المستوى الجزيئي منذ مليارات السنين، ومن بين مكتشفاته ثمَّة آلاف الأدوات الجزيئية المخصصة للخلايا لتستخدمها لمهام محددة. وتنتصب الهوائيات والكلّابات بين تلك الأدوات المخصصة لاستغلال قوانين الفيزياء والحساب.
فعلى سبيل المثال، إذا كنت خليَّةً بين عددٍ من الخلايا التي تحصل على بروتينٍ لاصق، فسرعان ما ستبدأ بتشكيل مداراتٍ متداخلة، فتأخذ الخلايا عالية الالتصاق موضعًا في الوسط بينما تتواجد الخلايا منخفضة الالتصاق في المحيط الخارجي. ولا يحتاج أحدٌ لتوجيه هذه العملية، إذ إنها ناجمةٌ عن القوانين الفيزيائية، بيدَ أنك لن تتمكن من الوصول إلى تلك الأحياء البنيوية ما لم تكن بحوزتك البروتينات الصحيحة لتستثمرها في هذا الخصوص. وإذا كنت جزءًا من كائنٍ متعدد الخلايا وكانت خلاياك تصنع جسيمات حديدية وترتبها على نحوٍ متراصف، فستجد نفسك فجأةً مشاركًا في قوة تسحبك إلى الشمال المغناطيسي. وقد كان الحقل المغناطيسي من قَبلُ غير مرئيّ لك، ولكن من خلال ترتيبك للبروتينات المناسبة بطريقةٍ معينة، ها أنت تحصل مجانًا على كفاءةٍ جديدة على حين غِرّة. وإذا ما اكتشفت قناةً أيونية ستتمكّن فجأةً من المشاركة في جميع أنوع الديناميات الكهربائية. وإذا ما حصلت على النوع المناسب من القنوات الأيونية، فسيغدو بمقدورك فورًا الحصول على دارة تغذية راجعة تمنحك الذاكرة الكهروحيويَّة/ بينما ستمنحك قنواتٌ أخرى بوابة لا- NOT المنطقية وبوابة الاقتران- AND، وهذا يمكّنك من تشكيل أيّ نوع من الوظيفة البوليانية- Boolean التي توفّر لك طرقًا لاستثمار قوانين المنطق أو الحساب. هذه ليست بقوانين فيزيائية، وإنما هي قوانين رياضية، وأنت لست بحاجةٍ لفهمها كي تتمكّن من استثمارها، فكل ما تحتاجه هو تطوير بروتينات تتيح لأنظمتك الفرعية فرصة الاقتران بتلك الديناميات. يشبه الأمر الطرق العديدة للقيام بالحساب، من رسم الأرقام الرومانية بواسطةً عصىً في الرمال، وكتابة الأرقام العربية على الورقة بقلم رصاص، أو استخدام الخرزات على المحسبة، أو القلّابات- flip flops على جهاز الكمبيوتر. وبصرف النظر عن ماهية الوسيلة المادية، فإذا كانت قادرةً على الاحتفاظ بالنتائج “في الذاكرة”، ريثما يتواصل الحساب، فسيكون بمقدورها العثور على المعلومات التي تكتشفها الهوائيّات واستخدامها.
ينبغي أن تتمثَّل القطع بحدّ ذاتها الخبرة اللازمة للعثور على أمكنتها المناسبة والقيام بمهامها الصحيحة.
لاحظ كيف يكون “بمقدورك” أن تكون خليَّةً مفردة، أو كائنًا متعدد الخلايا، أو عضوًا، أو نسيجًا ضمن كائنٍ متعدد الخلايا، بينما تحافظ على كفاءاتك المعلوماتية المكوّنة من “البراغي والصامولات” الأساسية الخاصة ببنى معالجة المعلومات. وضمن هذا المنظور الذي حرصنا على أن يكون محدودًا، ليست الوسائط بحاجةٍ لأن تتحلّى بالوعي والفهم والعقل، ولكنها بحاجةٍ لأن تنتظم في بنى لاستثمار الأنساق الفيزيائية التي تمكّنها من استخدام المعلومات -تبعًا لقوانين الحساب- لتنفيذ المهام، بدءًا بالمهمة الجوهرية المتمثّلة في حفظ الذات التي لا تتطلب منها إمداد نفسها بالطاقة اللازمة للسيطرة على أدواتها فحسب، بل أن تكون قادرة على التكيّف مع بيئاتها المحليَّة بطرقٍ تقوي إمكاناتها.
إذا عدنا إلى السيارة ذاتية التجميع، فأين تكمن الخبرة اللازمة لهذا التجميع؟ في القطع دون أدنى شك، لا في جزيئات الماء المتقافزة تقافزًا عشوائيًّا بينما تخلو الصورة من أيّ عاملٍ آخر. فينبغي أن تتمثّل القطع بحد ذاتها الخبرة اللازمة للعثور على أمكنتها المناسبة والقيام بمهامها الصحيحة. ويجوز أن يتم التعامل معها بوصفها وسائط ذات كفاءات تغنيها عن الحاجة إلى الفهم. وكما أظهر داروين كيف يُستغنى عن العمل الفكري الإعجازي لـ “المصمم الذكي”؛ ليحلّ محله مجرد ترتيب تعاقبي مرجح لتريليونات النسخ المتباينة للكائنات الحيَّة التي صَمَّمَتْ تدريجيًّا -دون علمٍ أو نيّةٍ منها- جميع الترتيبات الأنيقة للطبيعة. وهكذا ينبغي لنا الآن أن نستعيض عن المجمِّع الذكي الذي اعتاد أن يقرأ دليل التعليمات الذي أعده “المصمم الذكي” إن وجد، بسيرورةٍ تصاعدية توزع كل الذكاء اللازم على خواص التصميم الجيدة- الكفاءات- على مئات المستويات.
لا تعرف الخلايا المخروطيّة في شبكية العين كيف تحوّل الفوتونات إلى نبضات عصبية فحسب، بل تعرف أيضًا كيف تتصل بالعصبونات المناسبة لتضعها في تماسٍ مباشر مع الشبكات العصبية التي تنجز الرؤية لدى الكائنات الحيَّة المبصرة. ولكنَّ معرفة الخليَّة المخروطية -خلافًا المجمِّع الذكي فالذي يقرأ كتيّب التعليمات ليعرف كيف يصِل الأسلاك بين المصباحين في مقدمة السيارة والمدخرة في مؤخرتها- تتسم بكونها حسيرةً للحد الأقصى، فبيئتها مجهرية، وهي بمثابة الحكيم المجنون الذي يتمتع ببعض المواهب المذهلة بيد أنه لا يدرك تلك المواهب على الإطلاق. ولكن بفضل التواصل- بإرسال الإشارات- مع الجيران، يمكن لتلك الخليَّة الإسهام بكفاءتها المحلية الخاصة في نظامٍ موزّع يتمتع بقدراتٍ موجهة بالاستناد إلى المعلومات ويتمتع بمجال طويل من حيث المكان والزمان.
مع ذلك، يبدو أنَّ ثمَّة مشكلةً جوهريةً في هذا الصدد؛ إذ يستند التطوّر إلى مبدأ الأنانية، فكيف يمكن لأنظمة حية معقدة أن تنفذ أهدافًا جماعيةً ستعمل لأجلها وحداتها الخلوية الثانوية؟ وكيف يمكن لهذا التعاون أن ينشأ من أفعال الوسائط المتكاثرة الأنانية؟ ولنتأمل فيما يلي أداة التفكير التي ما زالت تستخدم منذ عقود لتحليل هذا اللغز، أو معضلة السجين، كما ورد في كتاب دانييل دينيت: “فكرة داروين الخطيرة”، الصادر عام 1995:
“تمثِّل “معضلة السجين” المثال الأشهر في نظرية الألعاب، وهي لعبة يسيرة تمارسها أنت وشخص آخر على سبيل المثال، فيزجّ بكما في السجن ريثما تخضعان للمحاكمة (على خلفية تهمة ملفّقة)، ويعرض الادعاء على كليكما صفقةً خاصة بمعزلٍ عن الآخر، فإذا تحلّى كل منكما بالصمود ولم تدليا بأيّ اعتراف ولم يورّط أحدكما الآخر، فعندئذٍ سيحظى كل منكما بحكمٍ مخفّف؛ إذ لا يتمتع المتهم الشاهد بتلك القوة، وإذا ما اعترفت أنت وورطت المتهم الآخر الذي بقي صامدًا، فعندئذٍ سيُخلي سبيلك ويسجن هو مدى الحياة، وإذا اعترف كلاكما وورَّط كلّ منكما الآخر، فستعاقبان جميعًا بالسجن لمدةٍ متوسطة. أمَّا إذا صمدت أنت، واعترف السجين الآخر، فلا شك أنه سيطلق سراحه وتسجن أنت مدى الحياة. فما الذي عليك أن تفعله؟”.
إذا تمكّنتما جميعًا من الصمود، فلا شك أنَّ هذا سيكون خيرًا لكما من أن تعترفا كلاكما. إذن، أليس بمقدور كل منكما أن يتعهّد للآخر بالصمود؟ (يشار إلى خيار الصمود وفقًا للمصطلحات المعتمدة في لعبة “معضلة السجين” بخيار “التعاون”). لا شكّ أنكما تستطيعان التعهّد لبعضكما، ولكنَّ الغواية ستأتيك لاحقًا بنقض العهد والخروج من السجن وترك المتهم الآخر الساذج يقع في مشكلةٍ عويصة، سواءً أخذت بهذه الغواية أم لا. وما دامت هذه اللعبة تقوم على مبدأ التناظر، فسيخالج المتهمَ الآخر إذن إغواءُ نفسه بأن يخدعك لتكون أنت الساذج. أفتستطيع أن تتكلّ على وفاء الآخر بوعده مجازفًا بقضاء بقيّة عُمرك في السجن؟ لعلّك ستكون بأمانٍ أكبر في حال طعنت أنت به، أليس كذلك؟ فبتلك الطريقة ستتلافى أسوأ النتائج قاطبةً، لا، بل قد يطلق سراحك. ولا ريب أنَّ المتهم الآخر سيفكّر في هذا الأمر، ولذلك قد يتخذ آمن خيار بخيانتك، وفي هذه الحالة سيتوجب عليك أنت أيضًا خيانته تلافيًا للكارثة، ما لم يكن كلاكما تقيًّا تقى يجعله يفضل قضاء حياته في السجن ولا ينكث العهد، وعندها سينتهي بكما الحال إلى قضاء فترة حكمٍ متوسطة. ليتكما تقدران على تجاوز هذه الأفكار وتتعاونا.
وبصفةٍ خاصة، فلنتأمل معضلة سجين محددة الحيّز المكاني من خلال شبكة وسائط (خلايا) تمارس كل منها لعبة معضلة السجين ضد جيرانها. وتتمثّل الطريقة التقليدية للتفكير في هذا الصدد في أنَّ عدد الوسائط ثابت، وهي متميزةٍ بصورةٍ دائمة، بينما تجد أنَّ الشيء الوحيد الذي يتغيّر ويتطوّر هو السياسة التي يتبعها كل وسيط لاتخاذ القرار بالتعاون مع جيرانه أو خيانتهم. ولكن لنتخيّل أنَّ بروتينًا خاصًّا (كونيكسين) اُكتشف من خلال التطوّر، وهو يتيح للبيئتين الداخليّتين لخليّتين متجاورتين أن تتواصلا تواصلًا مباشرًا عن طريق ما يشبه النفق الذي يتيح للجزيئات الصغيرة الانتقال عبره. وهكذا، يتيح اكتشاف التطوّر لنوع البروتين المعنيّ للنظام الفرصة لاستثمار ديناميات هامة.
فتواصل الأجزاء الداخلية لخليّتين يضمن مشاركتهما مناصفةً فيما بينهما للعناصر الغذائية وإشارات المعلومات والسموم وغيرها بسرعةٍ كبيرة. وبصورةٍ حاسمة، يجسّد هذا الاندماج نوعًا مباشرًا من “المصير المشترك”، فكلَّما حدث لأحدٍ شقي الوسيط المركب خير أو شر، أَثَّرَ على الشقّ الآخر تأثيرًا سريعًا. وفي ظلّ هذه الظروف، لا يستطيع أيّ شقّ أن يخون الشقّ الآخر أو أن يتجاهل رسائله، فإنه لمن سوء التكيّف بالنسبة لأحد الشقّين؛ إذ يقوم بأيّ أمرٍ سيئٍ تجاه الشقّ الآخر ما داما يتشاركان قدرهما ونصيبهما من الحياة. وهكذا يغدو التعاون التام مضمونًا على خلفية استحالة الخيانة نتيجة إزالة الحدود بين الوسيطين المعنيين. ويكمن العامل الرئيس على هذا الصعيد في كون التعاون لا يتطلب أيّ تخفيضٍ في مستوى الأنانية، فكلا الوسيطين المعنيين يحتفظ بأنانيته كاملةً مثلما كانت عليه من قبل؛ إذ تصبو الوسائط دائمًا إلى أن تكون في الصدارة، بيدَ أنَّ حدود الصدارة والذات التي تزود عنها بكل ما تملك ربما توسَّعت جذريًّا لتشمل نسيجًا أو عضوًا كاملًا.
المستورقات كائنات تلعب دور البطولة على صعيد إعادة تجديد الأعضاء، فكلَّما تعرَّضت للبتر أعادت التشكيل بصورةٍ مطابقةٍ تمامًا من الناحية التشريحية.
تزيل هذه الشبكة الفيزيولوجية بصورةٍ جوهرية الحدود بين الوسائط الصغرى، مشكّلةً نوعًا من الوسيط الخارق الذي يصعب فيه الاحتفاظ بالهُويَّة الفردية للوسائط الأصلية. ولا شكّ أنَّ هذه الحدود ليست بالحدود التشريحية، وإنما هي حدود فيزيولوجية أو وظيفية، وهي تتولّى مسؤولية فرز المقصورات الحسابية التي تتدفق البيانات داخلها بحرية بمضامين كثيرة من منظور نظرية الألعاب. وهكذا، تغدو المعلومات (الذاكرة) موضوعًا للمشاركة بين المجموع، ولا شكّ أنَّ هذا هو السبب الرئيس للاتصال بجارك؛ إذ إنك ترث مجانًا خلاصة ما تعلَّمه والدروس التي استخلصها من تاريخه، ولهذا قد دفع هو ثمن ذلك في سياق الجهود الاستقلابية.
علاوةً على ذلك، ثمَّة شيءٌ آخر مذهل يحدث حين توصّل الخلايا شبكات تراسل الإشارات الداخلية بينها؛ إذ تتصاعد قيم الضبط الفيزيولوجية التي تمثل الأهداف الفطرية في الدارات الخلوية الاستتبابية، وتزداد عمليات القياس التي تتحرى الانحرافات عن المجال الصحيح، فهي تُقاسُ في المجمعات الخلويَّة الكبيرة بصورةٍ أساسية ضمن المكان (النسيج أو العضو)، والزمان (الذاكرة وقدرات التوقع الأعلى ما دامت الشبكة المركبة من خلايا كثيرة تتمتع بقدرةٍ حسابية أكبر جدًّا من القدرات الخلويَّة الفردية)، وهذا يعني أنَّ أهدافها -الحالات الفيزيوكيميائية التي تمثّل عناصر الجذب في حيّز الحالة الخاص بها- ترتقي من الأهداف الاستتبابية الفيزيولوجية الدقيقة الخاصة بكل الخلايا على حدة نحو الاستتباب التشريحي الأكبر للتجديد والتطوير، فيمكنه تحقيق القوام المستهدف الصحيح من جميع أنواع التكوينات البدائية الغريبة، رغم الصخب والإرباكات الخارجية الهائلة على طول الطريق (انظر: الشكل رقم “1” أدناه)، وهذا هو النظام الذي انضم لاحقًا إلى النشاط الأكبر والأسرع والموجّه للأدمغة مع تطوّر الوصلات العصبية الكهربائية القديمة (الوصلات العابرة للثغرات المستندة إلى الكونيكسين)، إلى وصلاتٍ عصبيةٍ كيميائية حديثة، ومع تحوّل الخلايا (التي كانت تستخدم تراسل الإشارات الفيزيولوجي لتنسيق التخلّق) إلى عصبونات نحيلة محسَّنة السرعة.
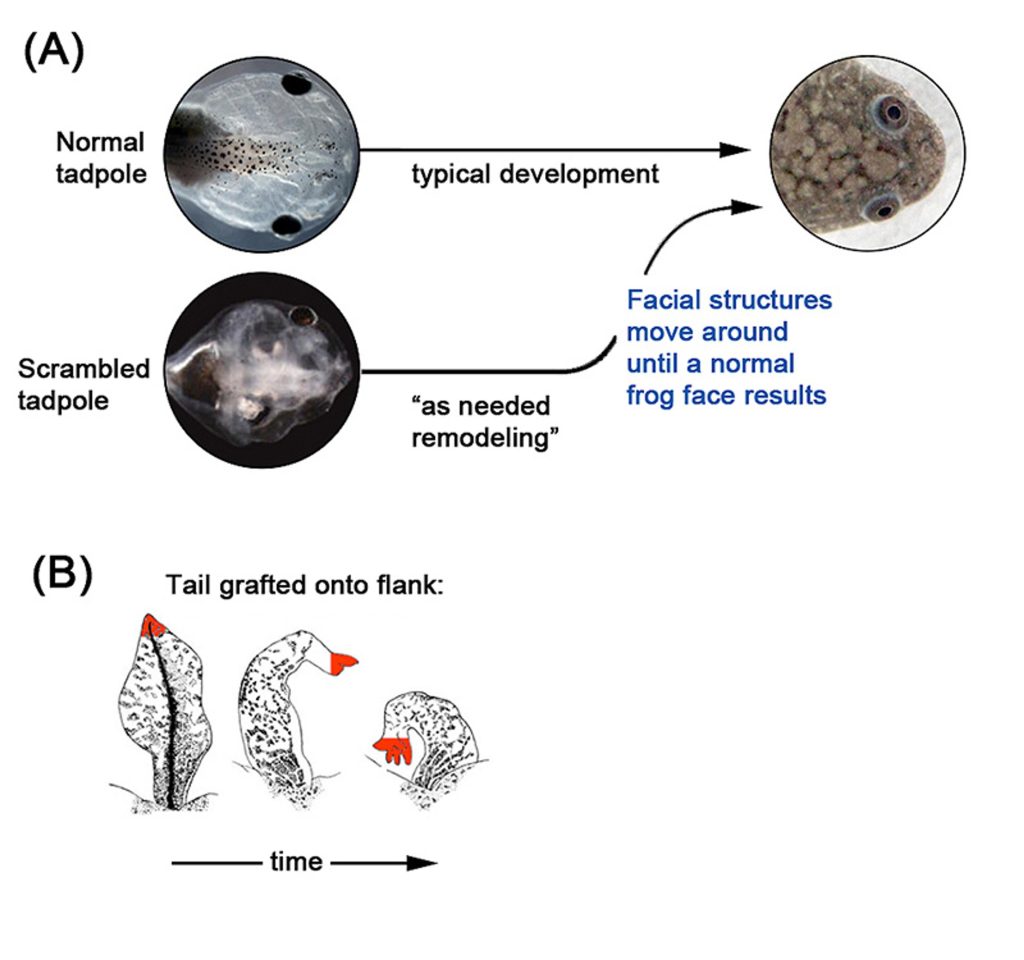
شرح الصورة1: تتعاون الخلايا بشكلٍ مرنٍ للوصول إلى بنى تشريحية معقدةٍ خاصة، كحال أعضاء وجه الشرغوف التي تعيد ترتيب نفسها إذا وُضِعَ في ظروفٍ غير طبيعية حتى تتمكن من تشكيل وجه ضفدعٍ طبيعي، أو مثلما يتحول ذيل السمندر الملتحم بخاصرة الحيوان إلى بنية أكثر ملائمة، أي حين يتحول الذيل إلى طرف. صورتا (الشرغوف الطبيعي) في اللوحة “أ” بواسطة دوغلاس جيه. بلاكيستون، مختبر ليفين لاب. وصورة الشرغوف المغير ترتيبه بواسطة فاندنبرغ وآخرون. أما الصور التي في اللوحة “ب” فهي بواسطة فارينيلا- فيروزا 1956م.
ثمَّة ارتباطٌ شديد بين مشكلة التعاون وبين مشكلة أصل العقول الموحدة المدمجة ضمن مجموعة (الخلايا، أو النمل، أو غيرها). والدينامية الرئيسة التي كشف التطوّر النَّقَّاب عنها هي نوعً من التواصل الذي يمنح الوسائط امتياز الوصول إلى منظومة المعلومات نفسها التي أتاحت المجال لقياس الذوات، وهذا ما شكّل نقطة الانطلاق للسلسلة المتصِلة من الوساطة المطردة. ولهذا أيضًا مؤداه الطبي، إذ أنَّ منع هذا التواصل الفيزيولوجي ضمن الجسد من خلال تعطيل عمل الوصلات العابرة للثغرات أو إدخال قطعٍ من البلاستيك بين الأنسجة بكل سهولة، هو ما يطلق العنان للسرطان، أيّ: الانعكاس الموضعي إلى حالة وحيد الخليَّة القديمة التي تغدو معها حدود الذات سطحَ الخليَّة المفردة وحده فيما بقيّة الجسم يغدو من منظورها مجرد “بيئة” صالحةٍ للاستثمار المحتكِر. وقد بتنا الآن نعلم أنَّ دفع الخلايا اصطناعيًّا للعودة إلى الاتصال الكهروحيويّ مع جيرانها، قد يؤدي إلى استرداد الحالة الطبيعية للخلايا السرطانية من خلال حثّها نحو الهدف الجماعي المتمثّل في حفظ النسيج وصونه.
ولهذا الرأي مؤدىً آخر بارز أيضًا يتمثّل في كون التعاون لا يتركز على الارتباطية الجينية بقدر ما يتعلّق بالتشغيل البيني الفيزيولوجي. فما دامت القطع الصلبة- Hardware تتمتع بالقدر الكافي من الجودة لتمكين التواصل الفيزيولوجي لهذا النوع، فلن تكتسي التفاصيل الدقيقة إذن الأهمية ذاتها التي تحظى بها الديناميات الاستتبابية متعددة النطاق مما تضمنه لها قوانين الفيزياء والحساب. فعلى سبيل المثال: بخلافنا نحن البشر الذين لا نورث أبناءنا التغييرات الأساسية الجارية في أجسامنا خلال مسيرة حياتنا، ديدان العريضات المستورقة غالبًا ما تتكاثر بالانشطار والتجديد، وهذا يعني أنها تمارس عملية التوريث الجسدي؛ إذ ينتقل كل تحوّل أساسي لا يقتل الخليَّة الجذعية إلى الجيل الجديد، إذ تتكاثر الخليَّة الجذعية للمساعدة على ملء النصف المفقود من الدودة. وتحمل خرائطها الجينية الدليل على تراكم مخلَّفاتٍ تعود لزمنٍ سحيق يربو على 400 مليون عام. لا بد أنها خرائط تعمّها الفوضى إذن. فما زلنا نفتقر لمعرفة التركيب الوراثي الصحيح لديدان دوجيسة جابونيكا- Dugesia japonica، ولذلك ثمَّة اختلاط في الصيغ الصبغية لهذه الديدان، لا، بل إنها لا تملك جميعًا عدد الصبغيات نفسه. ومع ذلك، فالمستورقات كائنات لها البطولة على صعيد إعادة تجديد الأعضاء، فكلَّما تعرَّضت للبتر أعادت التشكيل بصورةٍ مطابقةٍ تمامًا من الناحية التشريحية بدقةٍ متناهية بنسبة 100% رغم فوضى الخريطة الصبغية، وهذا ما يدعوه مهندسوّ الكمبيوتر باستقلالية القطع الصلبة، وإنه من الممكن ضمن مجال تحمّل واسع، تشغيلُ البرمجيات نفسها التي تبني الديدان على قطعٍ صلبة جزيئية مختلفة تمامًا؛ لأنَّ البرمجيات تتمتع بقوةٍ هائلة، وهي مصممة لبناء الناتج واسع النطاق ذاته مهما كلَّف الأمر (من حيث إبلاغ الخلايا المتنوعة بما ينبغي فعله لتحقيق ذلك).
كيف “تتبيّن” الخليَّة “المناسبة” هُويَّة مَن يجب عليها الوثوق به كشريك حياةٍ لها؟ فنحن لا نرى كل خليَّة تقيم الشراكة مع كل خليَّةٍ أخرى من جاراتها. ولذا يبدو أنَّ معضلة السجين ما زالت تطلّ برأسها، فهل ثمَّة نوع من سيرورة “التجربة والخطأ” الداروينية مما هو كامن في أفضل الوصلات العابرة للثغرات للالتزام بها؟ وكيف للخلايا الفردية أن تحدد الطريق التي يجب المضيّ منها؟ يمكن لأكثر الخلايا -على المستوى الخلوي- الاقتران بأكثر الخلايا الأخرى. وما يحدد كيفية حدوث ذلك هو البرمجيات الفيزيولوجية التي تحدد ما إذا كانت الوصلات العابرة للثغرات بين أيّ خليّتين متاحةً أم لا. وتجدر الإشارة إلى أنَّ البرمجيات الفيزيولوجية غير “مترابطة بالأسلاك”، وهي أيضًا ليست “برمجيات ثابتة”، فالوصلات العابرة للثغرات -شأنها شأن الوصلات العصبية- تتمتع بذاكرة وتتأثّر بالحالات السابقة للخلايا.
وكونها موصلات تيار ذات جهد كهربائي، فهي في واقع الأمر ترانزستورات. وإننا نعلم بالفعل مدى قوة الترانزستورات بوصفها عنصرًا أساسيًّا للذاكرة ودارات التغذية الراجعة (الاستتباب الأساسي أو التضخيم) وصناعة القرار المنسقة. للخلايا سياساتها الرامية إلى الارتباط بالخلايا الأخرى استنادًا إلى تشارك العناصر الغذائية وإضعاف السموم -وهي خدعة تعلّمتها من الأجسام الأوّليّة في الأغشية الحيويَّة للبكتريا-، فضلًا عن مشاركة المعلومات التي تمكّن من الارتقاء بمستوى التوقعات، ومن ثَمَّ إحراز النجاح الاستتبابي. وهي في واقع الأمر تتحدّ أمام أيّة معارضة ضمن سيرورة تعرف بالتطبيع، ومن ثَمَّ ستتعرض أيّة خليَّةٍ شاذة فيزيولوجيًّا لمحاولة جاراتها لإعادتها إلى جادة المخطط الفيزيولوجي واسع النطاق، إذ تحاول الخلايا المجاورة باختصارٍ تعديل جهدها عن طريق التوصيلات العابرة للثغرات مثلاً.
لعلَّها حكايةٌ ميكانيكية معقولة، ولكن ألا يتعلّق الحديث برمته حول الذاكرة وصنع القرار والتفضيلات والسلوك الموجّه بالتجسد فحسب؟ قد يودّ كثيرون التمسك برأيهم أنَّ المعرفة الحقيقية تكمن فيما يفعله الدماغ، وأمَّا ما يجري في الكيمياء الحيويَّة فيبدو أنه مجرد القيام بأمورٍ متماثلة. ولعلَّنا نقترح قلب هذه الفكرة المألوفة، فلا تكمن النيّة في تجسيد التكوينات الشكلية، بل في تطبيع المعرفة. ليس ثمَّة شيءٌ سحريّ مما يفعله الإنسان أو أيّ كائن حيّ ذكيّ آخر يفتقر للتاريخ التطوري، إذ يعني تناول التطور على محمل الجد طرح السؤال حول ما بدت عليه المعرفة طوال تاريخها. تحول البيانات الحديثة ضمن مجال المعرفة القاعدية دون إمكانية التمسك بتقسيم مصطنع للمعرفة “الحقيقية” ومعرفة “كأن- as if”، إذ إنَّ ثمَّة سلسلة متصلة نستطيع أن نحدد عليها مواقع جميع الأنظمة الحيَّة (وكثير من الأنظمة غير الحيَّة)، فيما يخصّ مقدار التفكير الذي قد ينتج عنها.
تكوّن الأنسجة والأعضاء والأدمغة والحيوانات والمجموعات مختلف أنواع العقول التي يمكنها إحراز أهداف أكبر.
بينما تتعلّق أكثر الحكايات شعبية حول كيفية تعاون الخلايا لتحقيق أهداف هائلة بالخلايا العصبية، ينبغي لك ألا تنسى أنَّ ثمَّة فارقًا جوهريًّا صغيرًا بين العصبونات وغيرها من الخلايا على اختلاف أنواعها. فإنه لمن المعلوم الآن أنَّ البروتينات المتصلة عصبيًّا والقنوات الأيونية والوصلات العابرة للثغرات على سبيل المثال، قد كانت بالفعل موجودةً لدى أسلافنا من وحيديّ الخليَّة، وكذا كانت تستخدمها الخلايا النشطة كهربائيُّا لتنسيق الأفعال في الحيّز التكويني التشريحي -إعادة النمذجة والتطور- قبل زمنٍ طويل من انضمامها إلى بعضها بغية إدارة نشاطٍ أسرع ضمن الحيز ثلاثي الأبعاد. وإذا كنت تؤيد أنَّ ثمَّة آلية يمكن من خلالها للخلايا النشطة كهربائيًّا أن تمثّل ذكريات الماضي، والحقائق المستقبلية المضادة، والأهداف واسعة النطاق، إذن فليس ثمَّة سببٌ يمنع الشبكات الكهربائية غير العصبية من تنفيذ نسخةٍ سهلة من الأمر عينه لإنجاز الاستتباب التشريحي. وقد أوضح علم تطور السلالات أنَّ العصبونات تطورت من أنواعٍ أبسط كثيرًا من الخلايا، وأنَّ بعض حيل الدماغ محسّنة السرعة قد اكتشفت في زمن الأغشية الحيويَّة البكتيرية على وجه التقريب، (ولعلَّ الحيلة الكبرى تتمثّل في التنامي نحو شبكات يمكنها أن تمثّل باطرادٍ حالاتٍ مستهدفةً أكبر وأوسع، وأن تنسق دارة “اختبر- شغِّل- اختبر- اخرج” عبر الأنسجة. إذن، ما زالت مسيرة المعرفة أشبه بالتسلّق البطيء البعيد كل البعد عن أيّ قفزةٍ سِحرية.
ولعلَّ النقطة المركزية في المنظومات الاستعرافية -وإن كان تنفيذها ماديًّا حتى الحيوانات والخلايا وأشكال الحياة المركبة والذكاء الاصطناعي والكائنات الفضائية المُحتملة- تتمثّل فيما تعرف كيف تكتشفه وتتمثّله كذاكرة، وتتوقعه، وتحدده، وبصورةٍ حاسمة تحاول أن تؤثّر عليه. ولندعُ هذا بالأفق الاستعرافي للنظم. ومن الطرق المُستخدمة في تصنيف النُظم الاستعرافية ومقارنتها -سواءً أكانت نظمًا استعرافية صناعية ومتطورة أو بسيطة أو معقدة- وضع الخرائط الخاصة بأحجام وأشكال الأهداف التي تستطيع أن تدعمها، فتتمثلها وتسعى إلى تحقيقها). ويتكوّن عقل كل وسيط من أحد أنواع الأشكال ضمن الحيّز الافتراضي للأحداث الماضية والمستقبلية الممكنة. وتتحدد درجة حيز هذا الشكل بالمدى الذي يمكن للوسيط خلاله أن يشعر بالأفعال وينفذها، فهل يعرف الأحداث ويتصرّف بغية التحكم بها ضمن مسافة قدرها سنتيمتر واحد أو لعلّها تمتد أمتارًا أو أميالًا؟ أمَّا البُعد المكاني، فيتحدد من خلال العمق الذي يمكن للوسيط أن يتذكّره في الماضي، والمدى الذي يمكن له أن يستشرفه في المستقبل، فهل يمكنه استهداف أمورٍ ستحدث بعد دقائق أم أيام أم عقود؟
لا شكّ أنَّ البشر يحظون بآفاقٍ استعرافية واسعة جدًّا، وهُم يعملون أحيانًا بجدٍ في سبيل أمورٍ ستحدث بعد فترة طويلة من رحيلهم وفي أماكن بعيدةٍ جدًّا. أمَّا الديدان فتصبو إلى أهدافٍ محلية فورية فقط. وما بين البشر والديدان، سنجد مجالًا واسعًا من الوسائط الأخرى الطبيعية والصناعية. وتمثّل هذه الطريقة في تحديد الأفق الاستعرافي للمنظومة نوعًا من التناظر البياني الزمكاني مع الطرق التي تمثّل فيها الفيزياء النسبوية مخروطًا ضوئيًّا للمراقب، بما معنَاه الحدود الأساسية لما يمكن لأيّ مراقب أن يتفاعل معه عن طريق التأثير أو المعلومات. ويُظهِر الشكل رقم “2” أدناه الأمثلة ذات الصِلة. يتعلّق الأمر برمته إذن بالأهداف، فالأهداف الاستتبابية للخلايا المفردة تتراوح حول حجم خليَّةٍ واحدة تقريبًا، ولديها ذاكرة محدودة وقدرة محدودة على التوقع أيضًا. تكوّن الأنسجة والأعضاء والأدمغة والحيوانات والمجموعات -كثيب النمل على سبيل المثال- مختلف أنواع العقول التي يمكنها تمثُّل أهداف أكبر وتذكُّرها وإحرازها. ويمكّننا هذا المخطط المفهومي من النظر في تفاصيل الماضي التي لا صِلة لها بما يخصّ المواد أو خلفية إنشائها، فضلًا عن التركيز على ما هو هام لتكوين وسيطٍ استعرافي على درجةٍ ما من التعقيد، أيّ: نطاق أهدافه. يمكن للوسائط أن تتآلف في شبكاتٍ مرتقيةً بأهدافها المحدودة والمحلية لتغدو أهدافًا أجلّ وأكثر مهابةً تنتمي إلى ذاتٍ موحدة أكبر حجمًا. ولا شك أنَّ أيَّ وسيط استعرافي قد يتكوّن من وسائط أصغر، لكل منها حدوده الخاصة بشأن حجم وتعقيد ما يصبو إليه ويعمل لأجله.
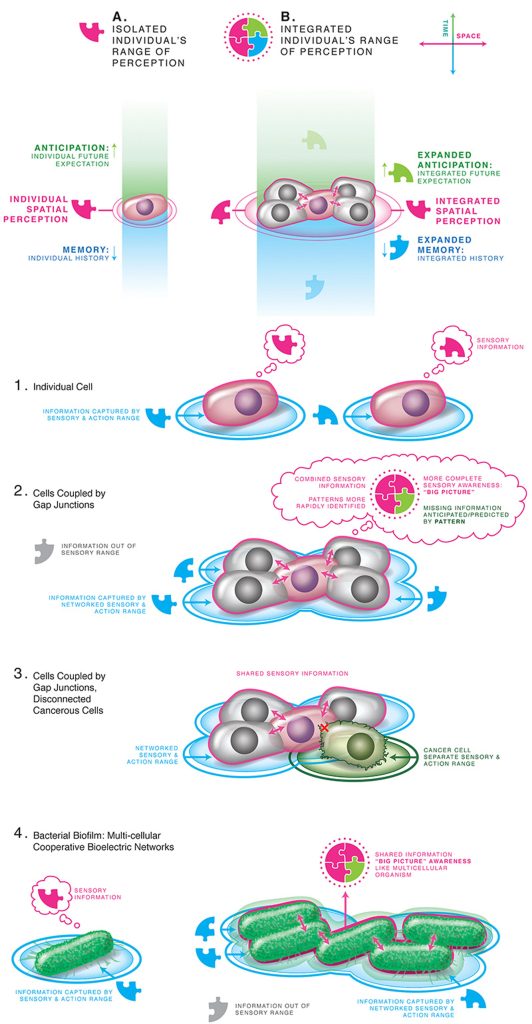
شرح الصورة 2: يجوز أن تتوسع الحدود الاستعرافية للذات من نطاقٍ زمكاني صغير لاستبباب فيزيولوجي أحادي الخلية إلى أهداف تشريحية أكبر لتخلّق الأعضاء من خلال ربط الخلايا عن طريق الوصلات العصبية الكهروحيوية لتكوّن منظومةً مفردة متماسكة. الرسم جيرمي غواي من مؤسسة بيرغرين كرييتيف انك.
إذا استندنا إلى هذا المنظور، نستطيع أن نتصوّر الإسهام الاستعرافي الدقيق للخليَّة المفردة في المشاريع الاستعرافية ومواهب مستكشفٍ يعمل على اكتشاف أراضٍ جديدة، فضلًا عن إسهامها أيضًا في قبيلة المُستكشف التي أمدّته بالتعليم والدعم، وذلك بفضل اللغة، وإسهامها -في نهاية المطاف- في فريقٍ من العلماء وغيرهم من المفكرين ممن يؤلّفون بين معارفهم وخبراتهم الاستكشافية، بفضل الأدوات الجديدة وإسهامها في الكون برمته، بل وفي الفضاءات المجرّدة الرياضية والشِّعرية والموسيقية. وبدلًا من التعامل مع “العبقرية” الإنسانية بصفتها أحد أشكال الصندوق الأسود المصنوع من مواد ذكيّة سِحرية، نستطيع أن نعيد تفسيرها على أنها توسعًا انفجاريًّا لحقيبةٍ من الحيل الميكانيكية، (ولكنها الاستعرافية المكتشفة بالاصطفاء الطبيعي على مدى مليارات السنين. ومن خلال توزيع الذكاء على الزمان -أو عصور التطوّر وسنوات التعلّم والنماء، وأجزاء الثواني الحسابية-، والمكان الذي لا يقتصر على الأدمغة والعصبونات الذكية فحسب، بل يشمل الأنسجة والخلايا الذكيّة والأنزيمات والجسيمات الريبية التي تتولَّى عملية التدقيق؛ سنصبح قادرين على توحيد ألغاز الحياة ضمن رؤية واحدةٍ تخطف الأنفاس.
————————————————————–
1- مايكل ليفين، رئيس قسم فانيفار بوش، وأستاذ متميز لعِلم الأحياء، في جامعة تافتس في ماساشوستس، ومدير مركز آلن للاكتشاف ومركز تافتس لعِلم الأحياء التجددي والنمائي.
2- دانييل دينت، أستاذ الفلسفة في قسم أوستن بي. فلتشر، والمدير المشارك لمركز الدراسات المعرفية في جامعة تافتس. وقد ألَّف ما يزيد على اثني عشر كتابًا، كان آخرها الكتاب الموسوم: “من البكتريا إلى باخ والعكس: تطوّر العقول”، الصادر عام 2017. وهو يعيش في ماساشوستس.
(وفق اتفاقية خاصة بين مؤسسة معنى الثقافية، ومجلة إيون).
تُرجمت هذه المقالة بدعم من مبادرة «ترجم»، إحدى مبادرات هيئة الأدب والنشر والترجمة.
الآراء والأفكار الواردة في المقالة تمثّل وِجهة نَظر المؤلف فقط.